Mechanisms of Allosteric Gene Regulation
TRAP ♦ Anti-TRAP ♦ SMK Riboswitch
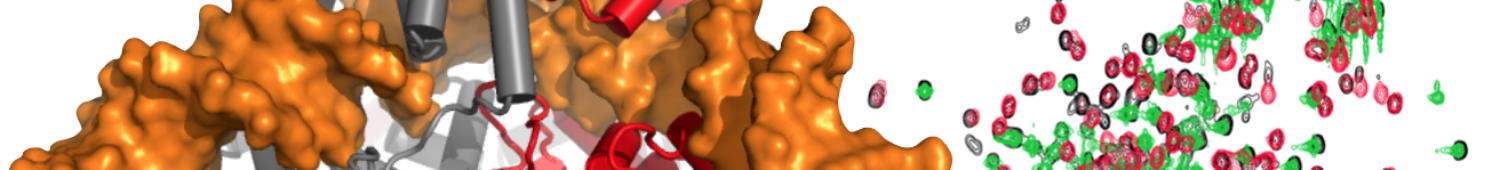
TRAP
The ring-forming oligomeric Bacillus trp RNA binding attenuation protein (TRAP), defines a paradigm for gene regulation by ligand-mediated alteration of the structure of non-coding RNA, and for mechanisms of both homotropic and heterotropic allostery. Undecameric (11-mer) TRAP serves as a sensor for intracellular tryptophan (Trp), which occupy its 11 identical sites, and thereby activates the protein for binding to specific RNA sequences in the 5′ untranslated regions of messenger RNAs. RNA binding by activated TRAP results in remodeling of RNA secondary structures that include (1) competing hairpins whose structure regulates transcription via aborted transcripts (termination), and (2) competing hairpins that regulate translation by alternately exposing or sequestering the ribosome binding site. The repressive RNA binding activity of Trp-activated TRAP can be blocked by the binding of another oligomeric protein, Anti-TRAP (AT), whose biosynthesis is regulated by sensing of the levels of uncharged tRNATrp. We seek to understand the role of protein dynamics communicating allosteric signals between Trp binding sites, and between Trp and its target RNA molecules.

Anti-TRAP
 Anti-TRAP (AT) exists in a pH-dependent equilibrium between a trimer and a tetramer of trimers, while only the trimer is able to inhibit RNA binding by TRAP. This implicates a new mechanism for pH-mediated regulation of gene expression via altered protein quaternary structure, and raises questions about mechanisms of pH sensing. Moreover, the polydentate structure of both AT3 and TRAP11 allow the interacting partners to assembly in heterogeneous chains involving multiple oligomers of each species. We aim to understand the roles of these phenomena in regulating expression of the Trp operon, and to explore the broader prevalence of these processes.
Anti-TRAP (AT) exists in a pH-dependent equilibrium between a trimer and a tetramer of trimers, while only the trimer is able to inhibit RNA binding by TRAP. This implicates a new mechanism for pH-mediated regulation of gene expression via altered protein quaternary structure, and raises questions about mechanisms of pH sensing. Moreover, the polydentate structure of both AT3 and TRAP11 allow the interacting partners to assembly in heterogeneous chains involving multiple oligomers of each species. We aim to understand the roles of these phenomena in regulating expression of the Trp operon, and to explore the broader prevalence of these processes.
SMK (SAM III) Riboswitch
 The SMK box riboswitch, which represents one of three known classes of S-adenosylmethionine (SAM)-responsive riboswitches, regulates gene expression in bacteria at the level of translation initiation. In contrast to most riboswitches, which contain separate domains responsible for ligand recognition and gene regulation, the ligand-binding and regulatory domains of the SMK box riboswitch are coincident. NMR experimentation within our lab provided direct evidence for a SAM-bound (BOUNDSMK) and an unbound conformation (ISOSMK). Indirect thermodynamic evidence supports the existence of an additional unbound conformation (PRIMEDSMK) that primes the riboswitch for binding SAM and occurs in a 5:1 exchange with the ISOSMK conformation. The existence of two unbound isomers experiencing such an exchange process makes SMK an ideal model for studying a pathway for spontaneous RNA conformational interchange. In collaboration with the Tina Henkin lab, we utilize a combination of NMR spectroscopy and isothermal calorimetry (ITC) to explore and characterize the free energy landscape of this riboswitch, elucidating the pathway that dictates the exchange between unbound conformations with opposing functions.
The SMK box riboswitch, which represents one of three known classes of S-adenosylmethionine (SAM)-responsive riboswitches, regulates gene expression in bacteria at the level of translation initiation. In contrast to most riboswitches, which contain separate domains responsible for ligand recognition and gene regulation, the ligand-binding and regulatory domains of the SMK box riboswitch are coincident. NMR experimentation within our lab provided direct evidence for a SAM-bound (BOUNDSMK) and an unbound conformation (ISOSMK). Indirect thermodynamic evidence supports the existence of an additional unbound conformation (PRIMEDSMK) that primes the riboswitch for binding SAM and occurs in a 5:1 exchange with the ISOSMK conformation. The existence of two unbound isomers experiencing such an exchange process makes SMK an ideal model for studying a pathway for spontaneous RNA conformational interchange. In collaboration with the Tina Henkin lab, we utilize a combination of NMR spectroscopy and isothermal calorimetry (ITC) to explore and characterize the free energy landscape of this riboswitch, elucidating the pathway that dictates the exchange between unbound conformations with opposing functions.